1O. DECAPOD BURROWS (THALASSINOIDES, OPHIOMORPHA) AND CRUSTACEAN REMAINS IN THE CALCARI GRIGI, LOWER JURASSIC, TRENTO PLATFORM (ITALY)
---------
Paolo Monaco
( 1st Workshop on Mesozoic and Tertiary decapod crustaceans, Montecchio Maggiore, Vicenza, October 6-8, 2000,)
contributions from: Alessandro Garassino (Museo Civico di Milano), Alice Giannetti, Paola Gigli, Silvia Roberti, Remo Ricco, Rita Umbrico (Perugia)
• MONACO P. (2000). Decapod burrows (Thalassinoides, Ophiomorpha) and crustacean remains in the Calcari Grigi, lower Jurassic, Trento platform (Italy). 1st Workshop on Mesozoic and Tertiary decapod crustaceans, Studi e Ricerche, Associazione Amici del Museo Civico “G.Zannato” Montecchio Maggiore (Vicenza), October 6-8, 2000, pp.55-57.
• MONACO P. & GARASSINO A. (2001). Burrowing and carapace remains of crustacean decapods in the Calcari Grigi, Early Jurassic, Trento platform. Geobios, 34 (3), 291-301.
© and permission of all colour photographs, text and charts by P. Monaco e-mail: pmonaco@unipg.it
Click to enlarge
|
|
|
|
|
 |
 |
 |
 |
Click on thumbnails to enlarge
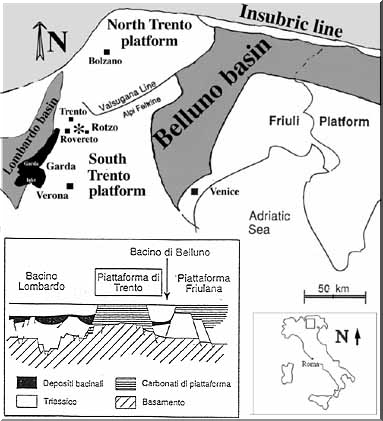
Burrows of decapod crustaceans are well preserved in the Pliensbachian of Trento carbonate platform, northern Italy (Fig.1). They are represented by several ichnoassemblages referred to Thalassinoides and Ophiomorpha isp., and are preserved as complex burrow systems from 2 to 16 cm in diameter. These burrows, still under detailed study, constitute complex boxworks, mazes, turning chambers at nodes and branched tunnel and reflect an intense burrowing activity of crustacean decapods forming horizontal and vertical patterns in the micritic substrate. Burrowing involve skeletal debris (Monaco 2000) as gastropods, bivalves, brachiopods, echinoderm, large foraminifera and so on, causing a reworking and transport deeply whithin the substrate of such skeletal particles, as indicated by peculiar taphonomic characteristics (Monaco 1999). The very abundant concentration of decapod crustacean burrows suggests an intense burrowing activity of such organisms during the Pliensbachian as indicated also by an exuvia of Phlyctisoma (Erymidae) as reported in Monaco and Garassino (in press), although we don’t know if this crustacean decapod was effectively an active tracemaker. Many shallowing-upwards parasequences occur and burrowing systems are concentrated mainly in deeper and quiet subtidal conditions (lower part). Conversely, in the middle part of cycles burrowing activity was progressively replaced by physical activity such as waves, or tidal-induced currents, storms and so on, and therefore burrow systems are reduced in size and intensity, especially in the upper part where intertidal/sopratidal conditions prevailed.
The studied area and methods
The Pliensbachian sequence of Calcari Grigi limestones, the so-called Rotzo member of the Trento carbonate platform of western Venetian Prealps (see geological setting and paleogeography in Bosellini & Broglio Loriga 1971; Clari 1975; Broglio Loriga & Neri 1976; Fugagnoli & Loriga Broglio 1996; Masetti et al. 1998 and references therein), have produced a large data set of taphonomic characteristics (Brett, 1995; Fürsich & Oschmann, 1993) gathered in a database regarding hundreds of collected fossiliferous samples (Monaco 1999a,b, 2000). Among all four members of Calcari Grigi commonly bioturbated, only the Rotzo member reveal crustacean decapod burrows and such trace fossils are very well preserved. Recently, during the last field work in the Folgaria area an exuvia of crustacean decapod Erymidae (Phlyctisoma) has been found (Monaco and Garassino in press). Three bioturbated sections are considered for crustacean burrowing analysis: the SVB of 27.15m, PVB of 39.90m and RVB of 23m in thickness respectively.
The analysis contemplate a description of burrow systems both in the lower surface of beds and whithin massive limestones, considering three-dimensional boxworks, meandered (regular or irregular) mazes, shafts, tunnels, turning chambers, and mounds of mud. The vertical distribution of burrows allows to reconstruct several level grounds (the so-called “tiers”). The grain arrangement of skeletals or peloids is revealed commonly by rounded ichnofabrics due to crustacean decapod activity (see wall structures of Ophiomorpha or Thalassinoides in Frey et al. 1978). Wall-pelleted ichnofabrics, structure of the burrow lining and lithified calcareous walls displaying concentric layering (Frey et al. 1978; Anderson & Droser 1998) have been considered and scannered slab of rocks have been recorded in a dataset for quantitative analysis (see Monaco 1999b). If boxworks are preserved on slab rock surfaces, the orientation and shape of trace fossils have been considered.
Decapod crustaceans: traces and body remains in the Pliensbachian swallowing-upward cycles
Thalassinoides and Ophiomorpha are trace fossil generally attributed to the activity of crustaceans, mainly but not only decapods (Frey et al. 1978). In the upper part of Calcari Grigi of Folgaria area, Pliensbachian in age, Thalassinoides is very abundant and well preserved mainly concerning Thalassinoides suevicus (Fig.2). Thalassinoides traces consist of three-dimensional, Y-shaped burrow systems, showing smooth-walled branches and cylindrical tunnels essentially developed in horizontal surfaces. Three type of Thalassinoides have been documented in the middle-upper part of the Rotzo member: Thalassinoides isp., Thalassinoides suevicus (type I or smaller from 2 to 5 cm in diameter, type II or medium from 6 to 10 cm, and type III or the larger one from 10 to 16 cm, respectively, Fig.2), and Thalassinoides isp.B (Monaco & Garassino, in press). Thalassinoides ichnogenus is easily recognizable for their horizontal mazes and cylindrical tunnels, generally slightly curved in plane view and more or less regurarly branched (see detailed description in Monaco & Garassino, in press). In particular T. suevicus reveals Y-shaped branches commonly enlarged at the bifurcation points (cfr. Uchman 1998; Fürsich 1998) and both the roof and the floor are generally not mammalated (cfr. Frey et al. 1978). Close but outside bifurcations crinoidal fragments, encrusted and bored shells concentrations, and large foraminifers (Orbitopsella sp.) are present. Rounded slab rock sections of T. suevicus type III reveal concentric grain distribution inside the trace fossil with a well preserved circular lining made of pellettoidal grainstone and a mammellated burrow boundary. Interiorly, the lumen is preserved and suggests the activity of the crustacean as found for type III traces. Other Thalassinoides isp.B burrows are gently curved, forming very wide U-shape traces (simple cylindrical tube up to 80 cm in lenght and 5 to 10cm in diameter), without turning chambers (very rare bifurcation points). They could be produced by other crustaceans such as carnivorous predators, the stomatopods, commonly know as “mantis shrimps” due to the raptorial appendages that they use to efficiently capture and subdue the prey (Caldwell & Dingle, 1976; Braithwaite & Talbot 1972). As reported by Bromley (1996) mantis shrimps are active burrowers and they first appeared in the Jurassic (e.g. Squilla, Kuhn 1973).
Ophiomorpha isp. consists of lined pellettoidal burrows generally infilled by coarse-grained detritus. Burrow systems with the walls formed by ovoid or discoidal pellets characterize the lower part of the Rotzo member (transition of Upper member/Rotzo member) where ripple-laminated calcarenitic bars or small-scale trough cross-bedded are present (Monaco et al., in prep.). In these calcarenitic limestones traces are from 2 to 4 cm in size (Ophiomorpha isp. A) and are the smaller among all pellettoidal crustacean burrows, similar to the Ophiomorpha nodosa of literature (Frey et al., 1978). Another type of Ophiomorpha isp., the larger one among traces of Rotzo member (Ophiomorpha isp.B), is found mainly in the upper part of Pliensbachian, revealing large and complex burrows from 5 to 15 cm in diameter (Monaco & Garassino, in press). These large burrows form frequently mammalated hyporeliefs from 7 to 15cm in diameter and up to 8-10cm in height with pellettoidal wall linings and abundantly encrusted skeletal debris, benthic foraminifera (Orbitopsella) and crinoids. Such Y-shaped mammalated reliefs represent lowermost tiers of very complex boxwork systems indicating deep bioturbation of large unknown organisms (decapod crustaceans? fish?). Other rare traces are very similar to Ophiomorpha irregulaire forming sinous horizontal burrows with elongated and mastoid pelletted walls extensively reworked by small, secondary burrowers such as Chondrites isp. (see Monaco & Garassino, in press).
A body remain of a decapod crustacean but not very well preserved, probably an exuvia, has been found in a dark calcareous bed of the SVB section (Monaco & Garassino, in press). It can be ascribed to the family Erymidae Van Straelen 1924, of genus Phlyctisoma, because is the only genus with an antennal spine so developed (Garassino 1996). Analysis of the body remain reveals a preservation of the rostrum and the cephalic part of carapace. The rostrum is long with pointed distal extremity and probably with one suprarostral tooth in the median part and a median longitudinal ridge. The cephalic part of carapace preserves the strong and elongate antennal spine. Phlyctisoma was established in the last century for two macruran decapod crustaceans of Cambridge Greensand (Lower Cretaceous, England), ascribed to two different species, P. tuberculatum and P. granulatum, in order to distinguish these forms from the other Jurassic erymids. Such distinction was essentially based on the different path of the grooves and on kind of ornamentation of the carapace. Actually ten species are ascribed to this genus, distributed from the Sinemurian (Lower Jurassic) to the Upper Campanian-Lower Maastrichtian (Upper Cretaceous).
Concerning the shallowing-upward cycles, in the Pliensbachian of Trento platform, the semi-relief tunnels of Thalassinoides characterize the lowermost part of the shallowing-upward taphocycles; taphonomic analysis indicate subtidal conditions and development of repetitive tiered intervals reaching 50cm in thickness (basal part of cycles). Details of burrow-wall morphology (smooth surfaces) at the base of cycles are well visible in lower surfaces of tiers because argillaceous-marly contacts with limestones were enhanced during diagenesis (Monaco & Garassino, in press). Another characteristic of subtidal intervals close to the storm wave base level (SWB) is the infilling of abandoned tunnels of crustacean decapods by coarse-grained detritus transported by hurricanes (see tubular tempestites, Wanless et al. 1988; Monaco, 1999b; 2000).
The middle and the upper parts of a cycle, conversely, are progressively much less burrowed and, therefore, nodularity induced by Thalassinoides and Ophiomorpha decreases upwards. Layer surfaces, therefore, are sharp and no tiered mazes have been found. In this middle-upper part of the cycle physical deposits are commonly dominant and deposited in a water depth between storm and fair-weather wave base (upper subtidal – intertidal): we have found tempestites, combined-flow deposits (waves and current along-shore, see Monaco 1999b; 2000), winnowed beds induced by tidal currents and coarse-grained ripples very rich in reworked lagoonal organisms and skeletal fragments. Finally, the upper part of cycles (intertidal-supratidal) reveal clear emersions, locally testified by plant remains such as Equisetites and other diagnostic features such as polygons, dessiccation fractures and mudcracked layers (Monaco et al., in prep.).
• ANDERSON B.G. & DROSER M.L. 1998 - Ichnofabrics and geometric configurations of Ophiomorpha within a sequence stratigraphic framework: an example from the Upper Cretaceous US western interior. Sedimentology, 45: 379-396.
• BOSELLINI A. & BROGLIO LORIGA C. 1971 - I "Calcari grigi" di Rotzo (Giurassico inferiore, Altopiano di Asiago) e loro inquadramento nella paleogeografia e nella evoluzione tettono-sedimentaria delle Prealpi Venete. Ann. Univ. Ferrara s.9, 5 (1): 1-61.
• BRAITHWAITE C.J.R. & TALBOT M.R. 1972 - Crustacean burrows in the Seychelles, Indian ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol., 11: 265-285.
• BRETT C.E. 1995 - Sequence stratigraphy, biostratigraphy, and taphonomy in shallow marine environments. Palaios, 10: 597-616.
• BROGLIO LORIGA C. & NERI C. 1976 - Aspetti paleobiologici e paleogeografici della facies a "Lithiotis" (Giurese inf.). Riv. Ital. Paleont. Strat. Mi., 82 (4): 651-706.
• BROMLEY R.G. 1996 - Trace fossils. Biology, taphonomy and applications. 361p., Chapman & Hall Publ. London (second edition).
• CALDWELL R. & DINGLE H. 1976 - Stomatopods. Scientific American, 234 (1): 80-89.
• CLARI P.1975 - Caratteristiche sedimentologiche e paleontologiche di alcune sezioni dei Calcari Grigi del Veneto. Mem. Degli Istit. Geol. e Mineral. Dell’Univ. di Padova, 31: 1-63.
• FREY R.W., HOWARD J.D. & PRYOR W.A. 1978 - Ophiomorpha: its morphologic, taxonomic, and environmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol., 23: 199-229.
• FUGAGNOLI A. & LORIGA BROGLIO C. 1996 - Revised biostratigraphy of Lower Jurassic shallow water carbonates from the Venetian Prealps (Calcari Grigi, Trento Platform, Northern Italy). Studi Trentini di Sc. Naturali, Acta Geologica, Trento, 73: 35-73.
• FÜRSICH F.T. & OSCHMANN W. 1993 - Shell beds as tools in basin analysis: the Jurassic of Kachchh, western India. Journ. of the Geol. Soc. London, 150: 169-185.
• FÜRSICH F.T. 1998 - Environmental distribution of trace fossils in the Jurassic of Kachchh (Western India). Facies, 39: 243-272.
• GARASSINO A. 1996 - The family Erymidae Van Straelen, 1924 and the superfamily Glypheoidea Zittel, 1885 in the Sinemurian of Osteno in Lombardy (Crustacea, Decapoda). Atti Soc. it. Sci. nat. Museo civ. Stor. nat. Milano, Milano, 135: 333-373.
• KUHN O. 1973 - Die Tierwelt des Solnhofener schiefers. Die Neue Brehm-Bücherei, A. Ziemsen Verlag, Wittemberg Lutherstadt: 119 p.
• MASETTI D., CLAPS M., GIACOMETTI A., LODI P. & PIGNATTI P. 1998 - I Calcari Grigi della Piattaforma di Trento (Lias inferiore e medio, Prealpi Venete). Atti Ticinensi di Scienze della Terra, Univ. di Pavia, Ed. New Press, 40: 139-183.
• MONACO P. 1999 - Taphonomic characteristics and biological overprints in the Calcari Grigi, Pliensbachian, Trento platform (northern Italy). Riv.Mus. Civ. Sc. Nat. “E.Caffi” Bergamo, 20 (1999), ISSN 0393-8700, 137-138.
• MONACO P. 1999 - Computer database as a tool to investigate taphonomy and events in carbonate platform environments. In: A. Farinacci & A.R. Lord (Eds.), Depositional Episodes and Bioevents, Paleopelagos, Spec. Public. n°2 (1999): 105-122, Roma.
• MONACO P. 2000 - Biological and physical agents of shell concentrations of Lithiotis facies enhanced by microstratigraphy and taphonomy, Early Jurassic, Gray limestones Formation, Trento area (Northern Italy). In: Hall R.L. & Smith P.L. (eds.): Advances in Jurassic Research 2000, Proceedings of the Fifth International Symposium on the Jurassic System, Vancouver BC, GeoResearch Forum 6, Trans Tech Publications, Switzerland: 473-486.
• MONACO P. & GARASSINO A. (in stampa). Burrowing and carapace remains of crustacean decapods in the Calcari Grigi, Early Jurassic, Trento platform. Geobios (2000)
• UCHMANN A. 1998 - Taxonomy and ethology of Flysch trace fossils: revision of the Marian Ksiazkiewicz collection and studies of complementary material. Annales Societatis Geologorum Poloniae, 68: 105-218.
WANLESS H.R., TEDESCO L.P. & TYRRELL K.M. 1988 - Production of subtidal tubular and surficial tempestites by hurricane Kate, Caicos Platform, British West Indies. Journ. of Sediment. Petrol., 58 (4): 739-750.